Evidence for Evolution
Article objectives
You are probably aware that the concept of evolution still generates controversy today, despite its wide acceptance. In The Origin of the Species, Darwin mentioned humans only once, predicting, ”Light will be thrown on the origin of man and his history.” Nevertheless, some people immediately distorted its far-reaching message about the unity of life into nearsighted shorthand: humans “came from” monkeys (Figure 1).

Figure 1: In Darwin’s time and today, many people incorrectly believe that evolution means “humans come from monkeys.” This interpretation does not do justice to Darwin’s theory, which holds that all species share common ancestry.
Evolution relates all of life – not just humans and monkeys. Biological evolution, like all scientific theories, is much more than just an opinion or hypothesis, it is based on evidence.
In science, a theory is an explanation which ties together or unifies a large group of observations. Scientists accept theories if they have a great deal of supporting evidence. In The Origin of the Species, Darwin took the time to compile massive amounts of fossil and biological evidence to support his ideas of natural selection and descent from common ancestors. He clearly and effectively compared animal breeding (artificial selection), which was familiar to most people, and natural selection. Because Darwin provided so much evidence and used careful logic, most scientists readily accepted natural selection as a mechanism for change in species. Since Darwin’s time, additional fossil and biological data and new fields of biology such as genetics, molecular biology, and biogeography have dramatically confirmed evolution as a unifying theory – so much so that eminent biologist Theodosius Dobzhansky wrote that “Nothing in biology makes sense except in the light of evolution.”
The Fossil Record: Structural Changes Through Time
Few would argue that dinosaurs roamed Earth in the past, but no longer exist. The fossil record is a revealing window into species that lived long ago. Paleontologists have carefully analyzed the preserved remains and traces of animals, plants, and even microorganisms to reconstruct the history of life on Earth. Relative (rock layer position) and absolute (radioisotope) dating techniques allow geologists to sequence the fossils chronologically and provide a time scale. Geology also reveals the environmental conditions of past species.
For many reasons, the fossil record is not complete. Most organisms decomposed or were eaten by scavengers after death. Many species lacked hard parts, which are much more likely to fossilize. Some rocks and the fossils they contained have eroded and disappeared. Moreover, much of evolution happens in the small populations that survive changes in environmental conditions, so the chance that intermediates will fossilize is low. Nevertheless, the current record includes billions of fossils – over 300 million from Los Angeles’ LaBrea Tar Pits alone, and an estimated 800 billion in South Africa’s Beaufort Formation. Analysts have identified 250,000 species among these remains.
Although the fossil record is far more detailed today than in Darwin’s time, Darwin was able to use it as powerful evidence for natural selection and common descent. Throughout geological history, species that appear in an early rock layer disappear in a more recent layer. Darwin argued that a species’ appearance recorded its origin, and that its disappearance showed extinction. Moreover, he noted remarkable similarities among structures in differing species, supporting common ancestry. Finally, he could often correlate environmental conditions with structures, supporting his idea that natural selection led to adaptations which improved survival within certain habitats.
As an example, let’s analyze a relatively complete set of fossils which record the evolution of the modern horse. Figure 2 sequences five species which show major evolutionary changes. The oldest fossil shows a fox-sized animal with slender legs and nearly vertical digits: Hyracotherium bit and chewed soft leaves in wooded marshlands. Geology and paleontology suggest that the climate gradually dried, and grasslands slowly replaced the marshes. Mesohippus was taller, with fewer, stronger digits – better able to spot and run from predators, and thus more likely to survive and reproduce in the new grasslands. Merychippus was taller still, and kept only one, enlarged digit – a hoof to run fast on the hard ground. By Pliohippus time, molar teeth had widened and elongated to grind the tough grasses. These fossils show gradual structural changes which correspond to changes in the environment. They appear to show a smooth, linear path directed toward the “goal” of the modern horse, but this is deceiving. These five fossils are merely “snapshots” of a bushy family tree containing as many as 12 genera and several hundred species. Some transitions are smooth progressions; others are abrupt. Together, they support natural selection and descent with modification from common ancestors.

Figure 2: The fossil record for relatives of the modern horse is unusually complete, allowing us to select a few which show major change over time. These changes can be correlated with environmental changes, supporting the ideas of evolution and natural selection. However, the linear arrangement is misleading; addition of all known fossils would show a branching, bushy path of descent and common ancestry.
Comparative Anatomy and Embryology
The evidence Darwin presented in The Origin of Species included not only fossils but also detailed comparisons of living species at all life stages. Naturalists in Darwin’s time were experts in comparative anatomy – the study of the similarities and differences in organisms’ structures (body parts). At different times during his life, Darwin studied the comparative anatomy of closely related species of marine mammals, barnacles, orchids, insectivorous plants, and earthworms.

Figure 3: Darwin’s Theory of Evolution explains both the similarities and the differences among living things. All flowering plants share leaves, petals, stamens, and pistil, but orchids have parallel-veined leaves and flowers with lips and fused stamens and pistil, while sundews have leaves with branching veins and flowers with equal petals and separate stamens and pistil. The two species of orchid (A and B) share a recent common ancestor, whereas all three species share a more distant common ancestor.
Species which share many similarities are closely related by a relatively recent common ancestor. For example, all orchids share parallel-veined leaves, two-sided flowers with a “lip,” and small seeds (Figures 3A and 3B). Species which share fewer similarities, sharing only basic features, are related by relatively distant ancestor. The sundew, one of the insectivorous plants Darwin studied, shares leaves and petals with orchids, but the leaves are wide with branching veins and the flowers are radially symmetrical rather than two-sided (Figure 3C). The many species of orchids, then, share a recent common ancestor, but they also share a more distant ancestor with the sundew.
Homologous and Analogous Structures
Similarities can show two different kinds of relationships, both of which support evolution and natural selection.
(1) Similarities shared by closely related species (species who share many characteristics) are homologous, because the species have descended from a common ancestor which had that trait. Homologous structures may or may not serve the same function. Figure 4 shows the forelimbs of mammals, considered homologous because all mammals show the same basic pattern: a single proximal bone joins a pair of more distal bones, which connect to bones of the wrist, “hand,” and digits. With this basic pattern, bats build wings for their lives in the air, whales form fins for their lives in the sea, and horses, as we have seen, construct long, hoofed legs for speed on land. Therefore, homologous structures support common ancestry.

Figure 4: Homologous structures are similarities throughout a group of closely related species. The similar bone patterns in bat’s wings, dolphin’s flippers, and horse’s legs support their descent from a common mammalian ancestor.
(2) Similarities shared by distantly related species may have evolved separately because they live in similar habitats. These structures are analogous because they serve similar functions, but evolved independently. Figure 5 compares the wings of bats, bird, and pterosaurs. Bats evolved wings as mammals, pterosaurs as dinosaurs, and birds from a separate line of reptiles. Their wings are analogous structures, each of which evolved independently, but all of which suit a lifestyle in the air. Note that although the wings are analogous, their bones are homologous: all three share a common but more distant vertebrate ancestor, in which the basic forelimb pattern evolved. Because analogous structures are independent adaptations to a common environment, they support natural selection.

Figure 5: The wings of pterosaurs, bats, and birds illustrate both homologous and analogous structures. Similarities in the patterns of bones are due to descent from a common vertebrate (reptilian) ancestor, so they are homologous. However, the wings of each evolved independently, in response to similar environments, so they are analogous, and provide evidence for natural selection.
Embryology
Embryology is a branch of comparative anatomy which studies the development of vertebrate animals before birth or hatching. Like adults, embryos show similarities which can support common ancestry. For example, all vertebrate embryos have gill slits and tails, shown in Figure 6. The “gill slits” are not gills, however. They connect the throat to the outside early in development, but in many species, later close; only in fish and larval amphibians do they contribute to the development of gills. In mammals, the tissue between the first gill slits forms part of the lower jaw and the bones of the inner ear. The embryonic tail does not develop into a tail in all species; in humans, it is reduced during development to the coccyx, or tailbone. Similar structures during development support common ancestry.
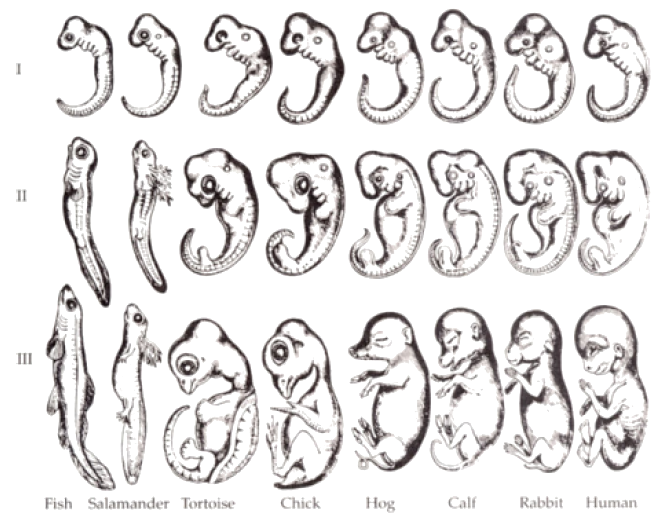
Figure 6: Comparative embryology reveals homologies which form during development but may later disappear. All vertebrate embryos develop tails, though adult humans retain only the coccyx. All vertebrate embryos show gill slits, though these develop into gill openings only in fish and larval amphibians. In humans, gills slits form the lower jaw and Eustachian tube. Many scientists consider developmental homologies evidence for ancestry, although some embryologists believe that these particular drawings exaggerate the similarities.
Vestigial Structures
Structures which are reduced and perhaps even nonfunctional, such as the human tail and the human appendix, are considered vestigial structures. The tail, of course, functions for balance in many mammals, and the human appendix may have served digestive functions in herbivorous ancestors. Whales, which evolved from land mammals, do not have legs or hair as adults; both begin to develop in embryos, but then recede. Vestigial leg bones remain, buried deep in their bodies, shown in Figure 7A.

Figure 7: Vestigial structures show evolutionary reduction or loss of unneeded structures which were useful to ancestors. A: Whales retain remnants of their mammalian ancestors’ leg bones (c). B: Cavefish lack the eyes and pigments important to their relatives who live in lighted habitats. C: True flies have reduced insects’ second pairs of wings to balancing knobs. D: We still show the reflex which raises hairs for insulation in cold air in our furry relatives, but all we have to show for our follicle’s efforts are goosebumps.
True flies have reduced the second pair of wings found in most insects to halteres for balance shown in Figure 7B. Cavefish lose both eyes and pigment, because both would require energy to build and are useless in the lightless habitat they have adopted shown in Figure 7C. You are probably very familiar with a fine example of a vestigial behavior: goosebumps raise the sparse hairs on your arms even though they are no longer sufficiently dense to insulate you from the cold by trapping warm air next to your skin; in most mammals, this reflex is still quite functional shown in Figure 7D. Most vestigial structures are homologous to similar, functioning structures in closely related species, and as such, support both common ancestry and (incomplete!) natural selection.
Molecular Biology
Did you know that your genes may be 50% the same as those of a banana?
Unknown in Darwin’s time, the “comparative anatomy” of the molecules which make up life has added an even more convincing set of homologies to the evidence for evolution. All living organisms have genes made of DNA. The order of nucleotides – As, Ts, Cs, and Gs - in each gene codes for a protein, which does the work or builds the structures of life. Proteins govern the traits chosen (or not) in natural selection. For all organisms, a single Genetic Code translates the sequence of nucleotides in a gene into a corresponding chain of 20 amino acids. By itself, the universality of DNA genes and their code for proteins is strong evidence for common ancestry. Yet there is more.
If we compare the sequence of nucleotides in the DNA of one organism to the sequence in another, we see remarkable similarities. For example, human DNA sequences are 98-99% the same as those of chimpanzees, and 50% the same as a banana’s! These similarities reflect similar metabolism. All organisms have genes for DNA replication, protein synthesis, and processes such as cellular respiration. Although metabolic processes do not leave fossils, similar DNA sequences among existing organisms provide excellent evidence for common ancestry.
The differences in DNA sequences are even more intriguing. Many are single base substitutions resulting from mutations accumulated through time. Assuming mutations occur randomly, the number of differences in bases between any two species measures the time elapsed since two organisms shared a common ancestor. This type of ”molecular clock” has confirmed traditional classification based on anatomy. Most scientists consider it sufficiently powerful to clarify or correct our understanding of evolutionary history. For example, human DNA differs 1.2% from chimpanzees, 1.6% from gorillas, and 6.6% from baboons; we can infer from this data that humans and chimpanzees share a relatively recent common ancestor, and that the common ancestor we share with gorillas lived much longer ago. Figure 8 shows a cladogram depicting hypothetical evolutionary relationships constructed with this data. Similarities and differences in the sequences of amino acids in proteins support common ancestry in the same way, because they are determined by DNA.

Figure 8: Cladograms use comparison data to construct diagrams showing evolutionary relationships. This cladogram uses comparisons of DNA nucleotide sequences to reveal patterns of descent from common ancestors. Molecular biology has supported and extended our understanding of evolutionary relationships based on traditional anatomy.
Heritability and variation in traits are essential parts of Darwin’s theory of evolution by natural selection. Since he published The Origin of the Species, rediscovery of Mendel’s identification of genes and how they are inherited has confirmed Darwin’s ideas. Molecular biology has clarified the nature of genes and the sources of variation. Comparative analysis of DNA and proteins continues to give us an exquisitely detailed view of patterns of variation, common ancestry, and how evolution works.
Biogeography
Australia, Africa, and South America occupy the same latitude, at least in part, and therefore have roughly the same climate. If plants and animals were distributed only according to their adaptations to habitat, we would expect the same species to occupy similar regions of these continents. However, the short-tailed monkeys, elephants, and lions in Africa differ significantly from the long-tailed monkeys, llamas, and jaguars of South America, and even more from the koalas, kangaroos, and Tasmanian devils of Australia. Biogeography studies the distribution of plants and animals and the processes that influence their distribution – including evolution and natural selection. Only geologic change and evolution can explain the distributions of many species, so biogeography is another kind of evidence for the theory of evolution.

Figure 9: Alfred Russel Wallace identified six major biogeographic regions: Nearctic, Neotropical, Palaearctic, Ethiopian, Oriental, and Australian Regions. Wallace explained the distributions of many animals and plants as a result of changes in geography and evolution.
Alfred Russel Wallace, who developed his own ideas of evolution and natural selection at the same time as Darwin, explained the distributions of many species in terms of changes in geography (such as formation of land bridges) and environment (for example, glaciations) and corresponding evolution of species. Figure 9 shows the six biogeographical regions he identified: Nearctic, Neotropical, Palaearctic, Ethiopian, Oriental, and Australian.

Figure 10: Biogeography explains the distribution of camel-like animals as a result of geographical changes and independent evolution. Today, the descendants of early camel ancestors are the dromedary in Africa, the Bactrian camel in Asia (center), and the guanaco (right) and llamas of South America.
Let’s consider just the camel family as an example, shown in Figure 10 of how biogeography explains the distribution of species. Fossils suggest that camel ancestors originated in North America. Distant fossils show structural similarities which suggest that their descendants migrated across the Bering land bridge to Asia and across the Isthmus of Panama into South America. These two isolated populations evolved in different directions due to differences in chance variations and habitat. Today’s descendants are llamas and guanacos in South America, and camels in Asia. Asian camels continued to migrate west into Africa, giving rise to two species – the dromedary in Africa, and the Bactrian in eastern Asia.

Figure 11: The locations of fossils such as Glossopteris on widely separated continents form contiguous patterns if the continents are joined. These patterns led to the theory of plate tectonics. Gondwanaland, a supercontinent of long ago, played an important part in evolution, natural selection and the history of life.
The distribution of some older fossils shows an opposite pattern; for example, fossils of a single species of fern, Glossopteris, have been found in South America, Africa, India, Antarctica, and Australia (Figure 11). Putting together many such distributions and a great deal of geologic data, Alfred Wegener showed that the continents were long ago united as Gondwanaland, and have since drifted apart. His theory of continental drift and its modern form, plate tectonics, help to further explain patterns of evolutionary descent in space and time.
Island Biogeography
Island biogeography studies archipelagos (oceanic island chains) as isolated sites for evolution. Both Darwin and Wallace used examples from isolated oceanic islands, such as the Galapagos and Hawaii, in their arguments for evolution and natural selection. Until humans arrived, terrestrial mammals and amphibians were completely absent on these islands. Darwin and Wallace showed that the animals and plants which were present had blown or drifted from one of the continents, or had descended – with modifications which suited the new habitats – from one of the original colonists. Terrestrial mammals and amphibians, having no powers of dispersal across oceans (until humans came along), were understandably absent.
Darwin's Finches
Only long after returning from his voyage did Darwin, with help from ornithologist John Gould, realize that the Galapagos birds he had collected but dismissed as uninteresting blackbirds, grosbeaks, finches, and a wren, were actually all closely related descendants of a single ancestral finch which had relatives on the South American mainland. Careful analysis showed that each of the 12 new species was confined and adapted to a specific habitat on a specific island. The finches, now known as “Darwin’s finches” (Figure 12A), clearly support both descent with modification and natural selection. Hawaiian honeycreepers (Figure 12B) are a more colorful but also more endangered example of the same evolutionary process of adaptive radiation. Bills ranging from thick and heavy (finch-like) for seed-eaters to long and curved for probing flowers illustrate the variations by which descendants of a single, original finch-like colonizer adapted to multiple ecological niches on the islands. Unfortunately, human destruction of habitat and introductions of rodents, the mongoose, and the mosquito which carries avian malaria have caused the extinction of 15 honeycreeper species, and still threaten the species which remain.
Figure 11: Darwin’s finches (above) on the Galapagos and honeycreepers (right) on Hawaii show the adaptive radiation of single finch ancestors which first colonized the islands. Each species show descent with modification, and the variety of bill shapes show adaptation to a specific niche. Many similar examples from island biogeography support evolution and natural selection. Honeycreepers are the finch-like palila (top right), the flower-probing I’iwi (center), and another nectar feeder, the amakihi (bottom).
Scientific Evidence
Altogether, the fossil record, homologies, analogies, vestigial structures, molecular uniformity and diversity, and biogeography provide powerful scientific evidence for the descent of today’s species from common ancestors. Some details of natural selection have been and are still being modified. However, the remarkable biological discoveries of the 150 years since Darwin published The Origin of the Species have dramatically strengthened support for his theory. Moreover, Darwin’s theory continues to enlighten new discoveries. Perhaps we could paraphrase Dobzhansky: Everything in biology makes sense in the light of evolution. The only piece still missing from the evidence puzzle is direct observation of the process itself. Darwin thought that humans could never witness evolution in action because of the vast time periods required. For once, however, he was mistaken.
Images courtesy of:
London Sketchbook. http://commons.wikimedia.org/wiki/Image:Caricatura_de_Darwin.jpg. Public Domain.
http://en.wikipedia.org/wiki/Image:Horseevolution.png. GNU-FDL.
http://commons.wikimedia.org http://commons.wikimedia.org/wiki/Image:Drosera_rotundifolia_chromolithograph.jpg/wiki/Image: Calypso_bulbosa_Nordens_Flora_416.jpg. Public Domain.
Jerry Crimson Mann. http://en.wikipedia.org/wiki/Image:Evolution_pl.png. Creative Commons.
John Romanes. http://commons.wikimedia.org/wiki/Image:Homology.jpg. Public Domain.
Romanes’s copy of Ernst Haeckel’s drawings. http://commons.wikimedia.org/wiki/Image:Haeckel_drawings.jpg. Public Domain.
http://commons.wikimedia.org/wiki/Image:Amblyopsis_spelaeus.jpg http://en.wikipedia.org/wiki/Image:Crane_fly_halteres.jpg http://commons.wikimedia.org/wiki/Image:Cold_urticaria.jpg. (d)Public Domain.
http://commons.wikimedia.org/wiki/Image:Lightmatter_chimp.jpg http://commons.wikimedia.org/wiki/File:Male_silverback_Gorilla.JPG http://commons.wikimedia.org/wiki/Image:Papio_ursinus.jpg. GNU-FDL, CC-BY-2.5, GNU-FDL, CC-BY-SA-2.5.
Alfred Russell Wallace. http://upload.wikimedia.org/wikipedia/commons/b/b1/Wallace_biogeography.jpg. Public Domain.
http://commons.wikimedia.org/wiki/Image:Bactrian.camel.sideon.arp.jpg http://commons.wikimedia.org/wiki/Image:Guanaco_09.24.jpg. GNU-FDL, GNU-FDL, Public Domain, Public Domain.
http://en.wikipedia.org/wiki/Image:Snider-Pellegrini_Wegener_fossil_map.gif. Public Domain.
http://commons.wikimedia.org/wiki/Image:Palila.jpg http://en.wikipedia.org/wiki/Image:Iiwi.jpg http://commons.wikimedia.org/wiki/Image:Amakihi.jpg. Public Domain, Public Domain, Public Domain, Public Domain.