Studying the History of Life
Article objectives
“There is grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one; and that, whilst this planet has gone cycling on according to the fixed law of gravity, from so simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved.” - Charles Darwin, Origin of Species. 1859
The history of life as we currently understand it is vast and wondrous and dramatic and humbling and ennobling. Vast is the almost unimaginable expanse of time during which life has flourished: four billion years is our current best estimate! Wondrous is the diversity of species throughout that time: some 30 million species occupy Earth today, and over 90% of all which have ever lived are extinct. Dramatic are the tales of change in environment and in diversity: ice ages, volcanism, continental drift, mass extinction, and bursts of evolutionary creativity have all shaped the natural environment. Humbling is the recognition that humans have played a relatively small part in the history; if the 4.6 billion-year story of Earth is reduced to a single cosmological day, humans occupy just the last minute and a half, and civilization covers less than the final second (Figure 1). Ennobling is the story’s revelation that we are related to and interdependent with all other species – back 4 billion years to “so simple a beginning” (Figure 2). Finally, the history of life suggests we might add one more striking impression: Terrifying is the realization that we are in many ways unique among species in our unprecedented power to change the environment, influence evolution, and destroy life’s diversity.

Figure 1: This Earth clock condenses the 4.5 billion years of earth’s history into a single 24-hour day. German names mark major geologic time periods. The last 17 seconds comprise the Quaternary period, spanning the past 2 million years. Human civilization appears only in the last second of the clock’s 24 hours.

Figure 2: A family tree of living things summarizes our understanding of the history of life and shows that humans and animals share common ancestors with all of modern life. This diagram demonstrates our current understanding of evolutionary relationships.
If we as a species occupy just the last minute and one-half of the cosmological day, how can we know the vast history of that 4.6 billion-year “day?” How did we arrive at 4.6 billion years as the age of the earth?
Tools: The Fossil Record, Aging the Ages, and Molecular Clocks
By age three, you probably knew that dinosaurs are part of the history of life. Our understanding of where they belong in the tale is relatively recent, but “dragon bones” have been known for thousands of years in China and Europe. Fossils are preserved remains or traces of organisms that provide extremely rare but vivid windows to the past. Because most parts of organisms decompose rapidly following death, fossilization is an exceptionally uncommon occurrence, and usually preserves only hard body parts, shown in Figure 3. Remains must be covered by sediment almost immediately. Buried organisms may experience mineralization (occasionally even within cells), or they may decay, leaving a space within the sediment later replaced with rock. Alternative pathways to fossilization include freezing, drying, trapping in resin (amber) or burial in anoxic (oxygen-free) environments. Trace fossils preserve footprints, burrows, droppings, eggs, nests, and other types of impressions. Overall, a great variety of types of fossils reveal the history of life, shown in Figure 4.

Figure 3: Scipionyx samniticus was a small dinosaur from Early Cretaceous Italy. This fossil of a juvenile only a few inches long is considered one of the most important vertebrate fossils ever discovered, because unlike most, it preserved internal organs as well as hard structures. Fossilization of an organism is itself a very rare event; preservation of soft tissues is even less likely.

Figure 4: Different types of fossils reveal the history of life. Clockwise from top left: Amber preserves an insect intact. Stone etches impressions of Edmontosaurus skin. Rock echoes a dinosaur’s footprint. Fossilized eggs recall a dinosaur of Mongolia. LaBrea Tar Pits fossilized the remains of a rich diversity of ice age animals. Permafrost preserved this female mammoth calf for nearly 10,000 years.
Images in rock tell us what kinds of organisms lived in the past, but the story of life cannot be told without knowing when various organisms appeared. Paleontologists use two methods to date fossils. The oldest method looks at position within a sedimentary column of rock to give a fossil’s relative age. If the rock layers are undisturbed, the deepest layers are the oldest, and layers near the surface are the youngest, shown in Figure 5. Widespread, short-lived index fossils can help identify rock layers of the same age spread around the earth, shown in Figure 6. Combining worldwide observations of relative position and composition resulted in a Geologic Time Scale for the Earth – a column of rock layers which reflects the history of sedimentary rock formation and changing life, stretching back to a time which apparently held no life. The fossil record showed patterns which, combined with his observations of living species, led Charles Darwin to conclude that all life on Earth descended from a single, simple common ancestor.

Figure 5: Relative aging dates sedimentary layers and the fossils they contain. Lower layers are older; upper layers are younger. Dinosaur fossils lie buried within this sedimentary formation in Green River, Utah.

Figure 6: Single-celled algae serve as index fossils to correlate rock layers located in different states. The middle-aged rock layer in South Carolina has apparently eroded from a similar deposit of sedimentary rock in Virginia. Careful worldwide studies of relative age by many geologists and paleontologists led to the Geologic Time Scale.
Relative age, however, only begins the story. Absolute aging, also known as absolute dating, uses radioactive isotopes, whose known half-lives can be used to calculate the number of years which have elapsed since a rock formed. Radioactive decay is a random but exponential process. An isotope’s half life gives the time period over which half of the material will decay to a different, relatively stable product, shown in Figure 7. The ratio of the original isotope to its decay product changes in a predictable way. This predictability allows the relative abundances of isotope and decay product to be used as a clock that measures the time from the incorporation of the isotope into a rock or a fossil to the present.

Figure 7: Exponential decay of a radioactive isotope such as carbon-14 occurs with a unique, predictable half-life (\(t^½\)) of 5,370 years. The amount of carbon-14 remaining in a fossil organism thus indicates the time elapsed since death, giving a measure of absolute age.
For example, half of a sample of Carbon-14 will decay to Nitrogen in 5,370 years. Cosmic rays cause the formation of Carbon-14 from the more common and stable Carbon-12 at a relatively constant rate, so carbon dioxide in the atmosphere contains relatively constant, predictable amounts. Organisms acquire carbon from various mechanisms – plants from \(CO_2\), and animals and other heterotrophs through food chains. When an organism dies, its carbon intake stops, and existing Carbon-14 atoms decay exponentially, according to their 5,370-year half-life. The proportion of Carbon-14 in the organism’s remains indicates the time lapsed since its death.
Carbon-14 has a relatively short half-life, so its use for absolute dating is limited to a maximum of about 60,000 years. Other isotopes are used to reach deeper into geological time. Uranium-238 and Uranium-235 decay to different isotopes of lead with half-lives of 4.46 billion and 704 million years, respectively, and together allow dating of rocks between 1 million and over 4.5 billion years old. Table 1 shows some of the many isotopes can be used to study rocks throughout Earth’s 4.6 billion year history.
Table 1: Isotopes Used to Measure Absolute Age of Rocks and Fossils
| Isotope | Decay Product | Half-life | Aging of Rocks or Fossils |
|---|---|---|---|
| Carbon-14 | Nitrogen | 5370 years | Up to 60,000 years |
| Uranium 238/235 | Thorium/Protactinium | 80,000/34,300 years | Hundreds of thousands of years |
| Potassium-40 | Argon | 1.3 billion years | Earth's oldest rocks |
| Uranium 238/235 | Lead | 4.5 billion/704 million years | 1 million to > 4.5 billion years |
Absolute aging techniques confirmed and brought into focus the rock layer story geologists and paleontologists had developed with relative dating. They pushed Earth’s history back 4.6 billion years, and showed that complex life evolved after some two billion years in which bacteria alone populated the Earth.
Further confirmation of common ancestry included molecular clocks, which measure changes in DNA or proteins to indicate degrees of relationship among species. Comparing DNA sequences of several species of primates, for example, shows that chimpanzees are more closely related to humans than are gorillas or baboons, shown in Table 2. If we assume uniform rates of mutation, we can estimate not only degree of relationship, but time back to common ancestry. Because DNA sequences (and mutations) determine the sequence of amino acids in proteins, Hemoglobin and other proteins are also used as “clocks.” Both DNA and protein clocks support a universal common ancestor for life, confirming the story which continues to unfold as new discoveries expand the fossil record. Molecular clocks, together with evidence from the fossil record, allows scientists to estimate how long ago various groups of organisms diverged evolutionarily from one another.
Table 2: DNA "Clock" Comparison of Primates
| Species | % Difference in Nucleotide Sequence, Compared to Humans |
|---|---|
| Human | 0 |
| Chimpanzee | 1.2 |
| Gorilla | 1.6 |
| Baboon | 6.6 |
A Geological Time Scale Measures the Evolution of Life
Observation of rock layers, dating techniques, and correlation of similar strata from around the world led to the development of a Geologic Time Scale (Figure 8). How does the scale divide 4.6 billion years of history? What themes emerge from its stories of the past?
One theme is almost unimaginable amounts of time. The deep time of Earth’s history is far beyond our experience, and our knowledge is far more detailed for recent millennia than for the distant past. A scale divided into evenly spaced periods of time would not show that detail. Instead, Geologic Time Scale divisions mark major events which highlight changes in climate, geography, atmosphere, and life. The largest units of time are Eons. Eons include smaller Eras, which in turn include Periods, Epochs, and Stages. Faunal stages identify specific fossil groups. Terms such as Upper/Late and Lower/Early divide parts of the scale into more recent and more distant subunits.
Four eons comprise the history of Earth, and their names refer to a second major theme of Earth history: the evolution of life. The Phanerozoic (“visible life”) Eon spans the most recent 545 million years and includes three Eras well known for their chronicle of life: the oldest Paleozoic, middle Mesozoic, and current Cenozoic. The Proterozoic (“before complex life”) Eon precedes the Phanerozoic, extending back 2.5 billion years. The Archean (“ancient”) and Hadean (“unseen”) Eons reach back to the formation of the Earth. The Precambrian Supereon combines the oldest three eons, and refers to the time before the first great explosion of life recorded in the fossil record - the Cambrian Period. The name “Cambrian” refers to Wales, where these fossils were first studied. Before this first period of the Phanerozoic, animals lacked hard body parts to contribute to the fossil record.

Figure 8: A linear arrangement of the Geologic Time Scale shows overall relationships between well-known time periods. Our knowledge of past life is concentrated in the most recent Eon, but the Phanerozoic occupies such a small proportion of the overall history of earth that eras, periods, and epochs are not precisely to scale. Relevant parts of the scale will show more detail with greater accuracy.
Patterns and Processes of Macroevolution
Throughout geologic time, the fossil record reveals dramatic changes in species and groups of species which have populated the Earth. Evolution at or above the species level is macroevolution, in contrast to microevolution, which describes changes within a species or population. Many scientists no longer emphasize the distinction, believing evolution to be a single process which includes both patterns. However, themes from the Geologic Time Scale illustrate macroevolution.
Although fossils dated back only to the Cambrian during Darwin’s time, radiometric dating has since identified fossil bacteria as old as the beginning of the Archean era 3.5 billion years ago. The geologic record shows over 2 billion years during which the only life was unicellular. The appearance of eukarytoic cells roughly 1.8 billion years ago marked a dramatic increase in cellular complexity. In rocks 1 billion years old, multicellular eukaryotes begin to appear, and by the end of the Precambrian, fossils record a variety of ancient multicellular organisms. The beginning of Cambrian Period marks an “explosion” of life, and in general, biodiversity has increased throughout the Phanerozoic, shown in Figure 9. Our current understanding of the fossil record confirms Darwin’s ideas that life began as tiny single-celled bacteria and over vast time evolved to produce the complexity and diversity we celebrate today.
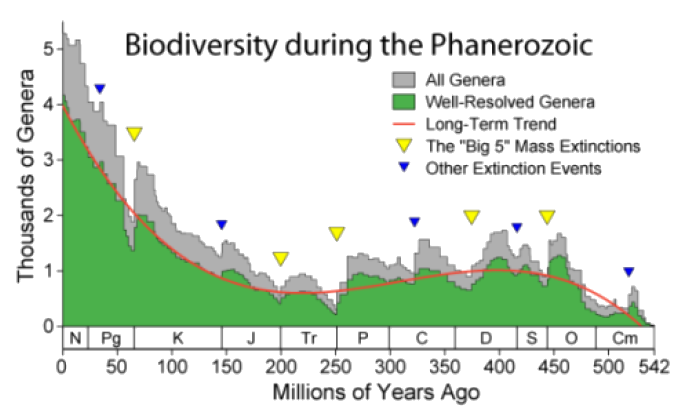
Figure 9: Estimates of numbers of marine genera throughout the last 542 million years support a gradual increase in biodiversity, interrupted by five major mass extinctions. Some scientists dispute the accuracy of such estimates, while others argue that they show regular cycles of extinction.
As recorded in fossils, the evolution of life was not smooth or steady. Mass extinctions and episodic speciation interrupted the overall pattern of increasing biodiversity, shown in Figure 9. These disruptions reflect dramatic changes in the environment of the Earth.
A major theme of the fossil record is loss of species. The death of a species – extinction – seems to be as much a characteristic of life as the death of individual organisms. Both seem closely linked to change in environment. Through mutation or sexual reproduction, offspring show variation. Individuals whose variations are not well suited to their environment die. Those whose variations are adaptive survive to reproduce. Death and differential reproduction result in adaptation to a changing environment.
These same forces of natural selection inevitably affect species: the fossil record indicates that up to 99.9% of all species that have ever lived on Earth are now extinct. Mass extinctions involve most major groups of organisms over a short period. The past 550 million years, when fossils are sufficiently abundant to tell a reliable story, show five mass extinctions in which more than 50% of animal species died. The most famous is the extinction which ended the reign of the dinosaurs 65 million years ago. Accelerated evolution may follow mass extinction, because the empty ecological niches make way for new species. After the non-birdlike dinosaurs disappeared, mammals rapidly evolved to fill the available niches. The fossil record shows numerous examples of episodic speciation, a pattern of periodic increase which includes these rebounds as well as bursts of evolution following major new “discoveries” or “ideas” – for example, the biochemical pathways for photosynthesis or cellular respiration.
Closely related to mass extinction is the theme of major environmental change throughout Earth’s history. Rock layers reflect critical changes in atmosphere and climate: oxidized iron deposits mark the introduction of oxygen gas to the atmosphere, and glacial deposits reflect periodic ice ages alternating with times of global warming. Craters and unique worldwide strata suggest that spectacular asteroid or comet collisions may have severely reduced solar radiation, and lava flows and ash suggests volcanism could have done the same. Massive geographic changes, now explained by plate tectonics theory, underlie volcanism as well as formation of new land bridges, seaways, and continents. Certain worldwide sedimentary deposits suggest significant sea level fluctuation, which may result from some of the aforementioned climate or plate tectonic changes. Life evolved against the backdrop of these often-catastrophic changes, and over 3.5 billion years of natural selection inevitably responded to them. Many of these changes are believed to have caused the mass extinctions and episodic speciation revealed in the fossil record.
Two caveats are critical in interpreting the history of life using the Geologic Time Scale. The first concerns the idea that evolution progresses via slow, steady, gradual change. We have already seen that mass extinction and episodic speciation interrupt the overall pattern of increasing biodiversity, but gradualism suggests that changes accumulate continuously as one species evolves to become another. An alternative, more recent theory, punctuated equilibrium, shown in Figure 10, proposes that species remain the same for long periods, and that change occurs infrequently but rather rapidly under unusual conditions such as geographic isolation or migration. The rather sudden appearance and disappearance of many individual species within the fossil record, noted even by Darwin, tends to support the latter theory. The idea of quantum evolution attempts to explain the origins of major groups (families, orders, and classes) as a response to drastic changes in environment or adaptive zones. The fossil record supports great variation in the rate of evolutionary change - from group to group and even among closely related lineages. Each of these ideas about pattern and rate may accurately describe one of many ways evolution works.

Figure 10: Two theories of evolutionary change - gradualism vs. punctuated equilibrium - are still debated. The former proposes continuous change, while the latter suggests that species remain constant for long periods of time and that change, when it occurs, is rapid.
A second caveat: the 4.6 billion year time scale makes it tempting to view evolution as linear, and perhaps even goal-directed. Time may be an arrow, but evolution is much more a bush of common ancestry–a family tree. Darwin recognized this - his sketch, shown in Figure 11, shows the pattern of speciation predicted by his theory of chance variation and adaptive selection. A very recent (August 2007) discovery encourages us to view our own human ancestry as a bush rather than a line. Radiometric dating of a new fossil of Homo habilis shows that this species coexisted with the ”more advanced” Homo erectus, shown in Figure 12. Previously, scientists considered the former an ancestor of the latter. The inappropriate expression “more advanced” implies the false, linear, goal-directed interpretation of evolution.

Figure 11: Two theories of evolutionary change - gradualism vs. punctuated equilibrium - are still debated. The former proposes continuous change, while the latter suggests that species remain constant for long periods of time and that change, when it occurs, is rapid.
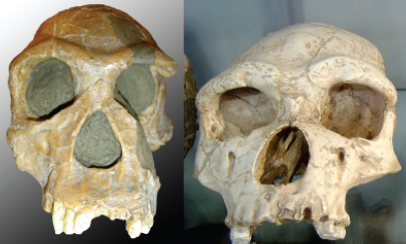
Figure 12: Homo habilis (left) was considered an ancestor to Homo erectus (right) until the 2007 discovery of a habilis fossil which showed that the two species coexisted. The history of the genus Homo, like the evolution of most species, is undoubtedly more bush-like than linear.
A famous example of “bushiness” in the history of life is adaptive radiation, a type of divergent evolution. This pattern of speciation involves the relatively rapid evolution from a single species to several species which fill a diversity of available ecological niches. Mass extinctions (the dinosaurs!), new volcanic islands (the Galapagos, or Hawaii), land bridge formation (the isthmus between North and South America) or “invention” of a new idea in evolution – all are events which “suddenly” open a variety of niches for adaptive radiation. In each case, a fundamental structure in one species is modified to serve new functions in different environments or modes of life. For example, forelimbs of mammals have become elongated with grasping hands for the forested habitats of monkeys, flattened into flippers for the aquatic habitats of whales, and spread into wings for the aerial habitats of bats, shown in Figure 13. Adaptive radiation explains – and the fossil record shows – that these groups all arose from one ancestor or a small group of common ancestors.
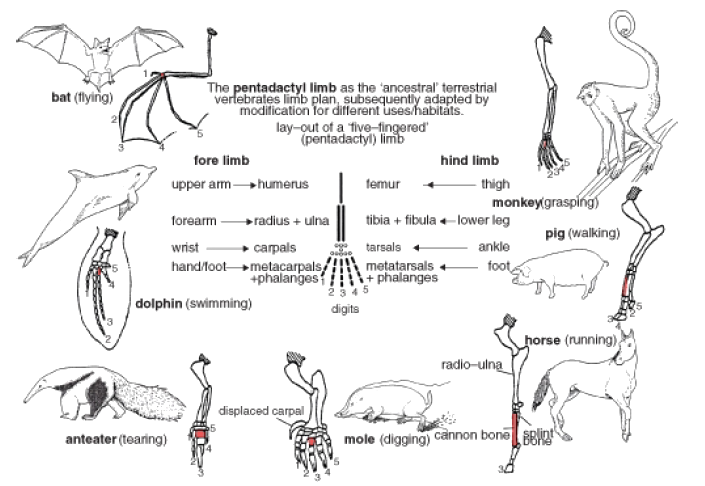
Figure 13: Forelimbs of mammals show adaptive radiation, or divergent evolution. Evolution has modified the original pattern in a common ancestor to suit a multitude of different environments.
In contrast to divergent evolution, whereby closely related species evolve different traits, convergent evolution involves distantly related species evolving similar traits. This pattern surfaces frequently in the history of life when different organisms occupy similar ecological niches. For example, three major groups of organisms have evolved wings for flight: reptiles (pterosaurs), birds, and mammals (bats), shown in Figure 14.

Figure 14: The wings of pterosaurs (1), bats (2) and birds (3) show convergent evolution. Similar structures adapt each group to flight, but each of the three types of wing evolved independently.
Australian fauna reveal both divergent and convergent patterns related to major geographical change, shown in Figure 15. Major groups of mammals evolved in the northern hemisphere and migrated to Australia across a land bridge. Later submerging of the land bridge isolated the Australian mammals, and the marsupials underwent their own adaptive radiation within their insular continent. Elsewhere, placental mammals evolved to out-compete the more primitive monotremes and marsupials, and underwent their own adaptive radiation. These independent radiations resulted in some wonderful examples of convergent evolution: An example: the marsupial Tasmanian wolf (now extinct) shared with the placental canines many adaptations to life as a hunting predator, shown in Figure 16.

Figure 15: Australia’s fauna demonstrates the importance of geography to evolution. Mammals evolved in the northern hemisphere and migrated to Australia across a land bridge (see A, above) which was later submerged (B). Marsupials persisted and underwent adaptive radiation in Australia. Elsewhere, the appearance of placental mammals spelled doom for the marsupials; placental mammals outcompeted them and underwent their own adaptive radiations. These separate radiations (green, orange, and red in B) resulted in a number of examples of convergent evolution.

Figure 16: The thylacine or Tasmanian wolf, a marsupial, closely resembles the golden jackal, a placental canine; both show similar adaptations to predatory life, demonstrating convergent evolution. Marsupials and placentals evolved independently due to the loss of a land bridge connecting Australia to southeast Asia, so they provide examples of convergent evolution.
One last, fascinating pattern within the history of life is coevolution. In coevolution, two species or groups of species influence each other’s evolution and therefore evolve in tandem. Relationships may be positive for one species or both, or an evolutionary arms race between predator and prey. Flowering plants depend on insects for pollination, so have evolved colors, shapes, scents, and even food supplies which are attractive to certain insect species. Insects, in turn, have evolved mouthparts, senses, and flight patterns which allow them to respond to and benefit from specific floral “offerings,” shown in Figure 17. The Endosymbiotic Theory describes a special form of co-evolution: Mitochondria and chloroplasts evolve within eukaryote cells, yet because these organelles have their own DNA sequence, different from that of the nucleus in the “host” cell, organelle and host cell evolve in tandem – each influences the evolution of the other.

Figure 17: Impressive proboscis and vivid colors! Hawk moths and the zinneas influence each other’s evolution, because the flower depends on the moth for pollination, and the moth feeds on the flower.
Closely related to coevolution is coextinction. If one member of a pair of interdependent species becomes extinct, the other is likely to follow. Famous examples were two species of bird lice which were obligate parasites on the passenger pigeon, shown in Figure 18. When “Martha,” a resident at the Cincinnati Zoo thought to be the last passenger pigeon in the world died on September 1, 1914, the extinction of the bird lice species followed. Alas, one louse species was later rediscovered on a band-tailed pigeon, and the other species had been misidentified.

Figure 18: The passenger pigeon and a parasitic species of louse (not the one pictured above) demonstrate coevolution and potential coextinction. Each species influenced the other’s evolution, and when the host became extinct in 1914, the parasite narrowly escaped extinction only because an alternate host – the band-tailed pigeon – survived.
Images courtesy of:
Hannes Grobe. http://commons.wikimedia.org/wiki/Image:Earth_clock_hg.png. CC-BY-SA 2.5.
http://commons.wikimedia.org/wiki/Image:Phylogenetic_tree.svg. Public Domain.
Giovanni Dall’Orto. http://commons.wikimedia.org/wiki/Image:9121_-Milano%2C_Museo_storia_naturale-Scipionyx_samniticus-_Foto_Giovanni_Dall%27Orto_22-Apr-2007a.jpg.
http://commons.wikimedia.org/wiki/Image:Edmontosaurusskin.jpg
http://en.wikipedia.org/wiki/Image:ClaytonLakeStateParkDinosaurFootprint.jpg
http://commons.wikimedia.org/wiki/File:Dinosauriereier_mongolei.JPG
http://commons.wikimedia.org/wiki/Image:Tar-bigpit.jpg
http://en.wikipedia.org/wiki/Image:Mammothzu2.jpg. Public Domain,Public Domain,GNU-FDL,CC-BY-SA 2.5
http://commons.wikimedia.org/wiki/Image:Green_River_UT_2005-10-14_2104.jpg. CC-BY-SA 2.5.
http://commons.wikimedia.org/wiki/Image:Biostratigraphie_usgs.png. Public Domain.
http://commons.wikimedia.org/wiki/Image:Exponential_Decay_of_Nuclei_with_Halflife-de.svg. GNU-FDL.
http://en.wikipedia.org/wiki/Image:Phanerozoic_Biodiversity.png. GNU-FDL.
http://en.wikipedia.org/wiki/Image:Darwins_first_tree.jpg. Public Domain.
http://en.wikipedia.org/wiki/Image:Punctuatedequilibrium.png. Public Domain.
http://commons.wikimedia.org/wiki/Image:Homo_erectus_tautavelensis.jpg. GNU-FDL,GNU-FDL.
http://en.wikipedia.org/wiki/Image:Evolution_pl.png. GNU-FDL.
John Romanes. http://commons.wikimedia.org/wiki/Image:Homology.jpg. Public Domain.
CK-12 Foundation. http://commons.wikimedia.org/wiki/Image:Evolution_con_dis.png. GNU-FDL.
http://commons.wikimedia.org/wiki/File:Golden_Jackal_sa02.jpg. Public Domain,Public Domain.
http://commons.wikimedia.org/wiki/Image:Macroglossum_stellatarum.jpg. GNU-FDL.
http://commons.wikimedia.org/wiki/Image:Louse_diagram%2C_Micrographia%2C_Robert_Hooke%2C_1667.jpg. Public Domain,Public Domain.