Cell Transport and Homeostasis
Article objectives
Probably the most important feature of a cell’s phospholipid membranes is that they are selectively permeable. A membrane that is selectively permeable has control over what molecules or ions can enter or leave the cell, as shown in Figure 1. The permeability of a membrane is dependent on the organization and characteristics of the membrane lipids and proteins. In this way, cell membranes help maintain a state of homeostasis within cells (and tissues, organs, and organ systems) so that an organism can stay alive and healthy.

Figure 1: A selectively permeable membrane allows certain molecules through, but not others.
Transport Across Membranes
The molecular make-up of the phospholipid bilayer limits the types of molecules that can pass through it. For example, hydrophobic (water-hating) molecules, such as carbon dioxide (\(CO_2\)) and oxygen (\(O_2\)), can easily pass through the lipid bilayer, but ions such as calcium (\(Ca^{2+}\)) and polar molecules such as water (\(H_2 O\)) cannot. The hydrophobic interior of the phospholipid does not allow ions or polar molecules through because they are hydrophilic, or water loving. In addition, large molecules such as sugars and proteins are too big to pass through the bilayer. Transport proteins within the membrane allow these molecules to cross the membrane into or out of the cell. This way, polar molecules avoid contact with the nonpolar interior of the membrane, and large molecules are moved through large pores.
Every cell is contained within a membrane punctuated with transport proteins that act as channels or pumps to let in or force out certain molecules. The purpose of the transport proteins is to protect the cell’s internal environment and to keep its balance of salts, nutrients, and proteins within a range that keeps the cell and the organism alive.
There are three main ways that molecules can pass through a phospholipid membrane. The first way requires no energy input by the cell and is called passive transport. The second way requires that the cell uses energy to pull in or pump out certain molecules and ions and is called active transport. The third way is through vesicle transport, in which large molecules are moved across the membrane in bubble-like sacks that are made from pieces of the membrane.
Passive Transport
Passive transport is a way that small molecules or ions move across the cell membrane without input of energy by the cell. The three main kinds of passive transport are diffusion, osmosis, and facilitated diffusion.
Diffusion
Diffusion is the movement of molecules from an area of high concentration of the molecules to an area with a lower concentration. The difference in the concentrations of the molecules in the two areas is called the concentration gradient. Diffusion will continue until this gradient has been eliminated. Since diffusion moves materials from an area of higher concentration to the lower, it is described as moving solutes ”down the concentration gradient.” The end result of diffusion is an equal concentration, or equilibrium, of molecules on both sides of the membrane.
If a molecule can pass freely through a cell membrane, it will cross the membrane by diffusion (Figure 2).
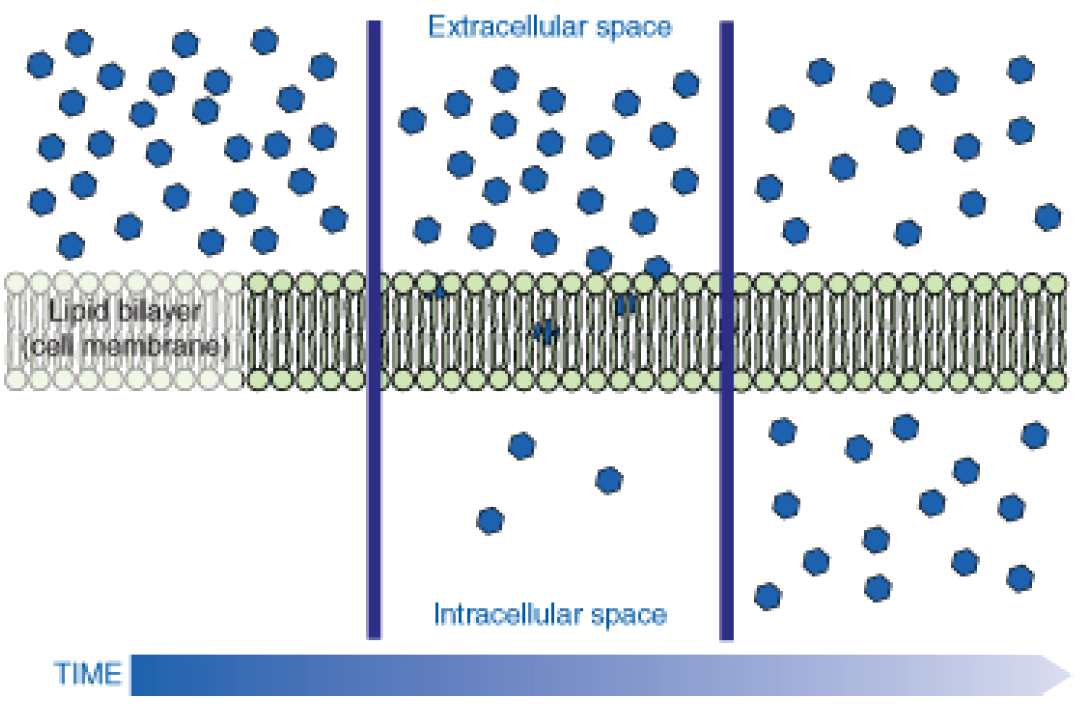
Figure 2: Molecules move from an area of high concentration to an area of lower concentration until an equilibrium is met. The molecules continue to cross the membrane at equilibrium, but at equal rates in both directions.
Osmosis
Imagine you have a cup that has 100ml water, and you add 15g of table sugar to the water. The sugar dissolves and the mixture that is now in the cup is made up of a solute (the sugar), that is dissolved in the solvent (the water). The mixture of a solute in a solvent is called a solution.
Imagine now that you have a second cup with 100ml of water, and you add 45 grams of table sugar to the water. Just like the first cup, the sugar is the solute, and the water is the solvent. But now you have two mixtures of different solute concentrations. In comparing two solutions of unequal solute concentration, the solution with the higher solute concentration is hypertonic, and the solution with the lower concentration is hypotonic. Solutions of equal solute concentration are isotonic. The first sugar solution is hypotonic to the second solution. The second sugar solution is hypertonic to the first.
You now add the two solutions to a beaker that has been divided by a selectively permeable membrane. The pores in the membrane are too small for the sugar molecules to pass through, but are big enough for the water molecules to pass through. The hypertonic solution is on one side of the membrane and the hypotonic solution on the other. The hypertonic solution has a lower water concentration than the hypotonic solution, so a concentration gradient of water now exists across the membrane. Water molecules will move from the side of higher water concentration to the side of lower concentration until both solutions are isotonic.
Osmosis is the diffusion of water molecules across a selectively permeable membrane from an area of higher concentration to an area of lower concentration. Water moves into and out of cells by osmosis. If a cell is in a hypertonic solution, the solution has a lower water concentration than the cell cytosol does, and water moves out of the cell until both solutions are isotonic. Cells placed in a hypotonic solution will take in water across their membrane until both the external solution and the cytosol are isotonic.
A cell that does not have a rigid cell wall (such as a red blood cell), will swell and lyse (burst) when placed in a hypotonic solution. Cells with a cell wall will swell when placed in a hypotonic solution, but once the cell is turgid (firm), the tough cell wall prevents any more water from entering the cell. When placed in a hypertonic solution, a cell without a cell wall will lose water to the environment, shrivel, and probably die. In a hypertonic solution, a cell with a cell wall will lose water too. The plasma membrane pulls away from the cell wall as it shrivels. The cell becomes plasmolyzed. Animal cells tend to do best in an isotonic environment, plant cells tend to do best in a hypotonic environment. This is demonstrated in Figure 3.
When water moves into a cell by osmosis, osmotic pressure may build up inside the cell. If a cell has a cell wall, the wall helps maintain the cell’s water balance. Osmotic pressure is the main cause of support in many plants. When a plant cell is in a hypotonic environment, the osmotic entry of water raises the turgor pressure exerted against the cell wall until the pressure prevents more water from coming into the cell. At this point the plant cell is turgid.
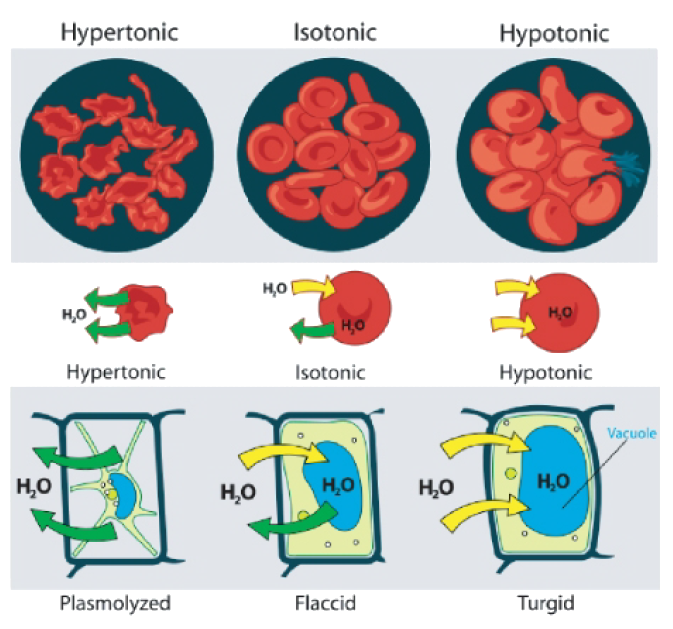
Figure 3: Unless an animal cell (such as the red blood cell in the top panel) has an adaptation that allows it to alter the osmotic uptake of water, it will lose too much water and shrivel up in a hypertonic environment. If placed in a hypotonic solution, water molecules will enter the cell causing it to swell and burst. Plant cells (bottom panel) become plasmolyzed in a hypertonic solution, but tend to do best in a hypotonic environment. Water is stored in the central vacuole of the plant cell.
The effects of osmotic pressures on plant cells are shown in Figure 4.

Figure 4: The central vacuoles of the plant cells in the left image are full of water, so the cells are turgid. The plant cells in the right image have been exposed to a hypertonic solution; water has left the central vacuole and the cells have become plasmolysed.
Osmosis can be seen very effectively when potato slices are added to a high concentration of salt solution (hypertonic). The water from inside the potato moves out of the potato cells to the salt solution, which causes the potato cells to lose turgor pressure. The more concentrated the salt solution, the greater the difference in the size and weight of the potato slice after plasmolysis.
The action of osmosis can be very harmful to organisms, especially ones without cell walls. For example, if a saltwater fish (whose cells are isotonic with seawater), is placed in fresh water, its cells will take on excess water, lyse, and the fish will die. Another example of a harmful osmotic effect is the use of table salt to kill slugs and snails.
Controlling Osmosis
Organisms that live in a hypotonic environment such as freshwater, need a way to prevent their cells from taking in too much water by osmosis. A contractile vacuole is a type of vacuole that removes excess water from a cell. Freshwater protists, such as the paramecia shown in Figure 5, have a contractile vacuole. The vacuole is surrounded by several canals, which absorb water by osmosis from the cytoplasm. After the canals fill with water, the water is pumped into the vacuole. When the vacuole is full, it pushes the water out of the cell through a pore. Other protists, such as members of the genus Amoeba, have contractile vacuoles that move to the surface of the cell when full and release the water into the environment.

Figure 5: The contractile vacuole is the star-like structure within the paramecia (at center-right)
Facilitated Diffusion
Facilitated diffusion is the diffusion of solutes through transport proteins in the plasma membrane. Facilitated diffusion is a type of passive transport. Even though facilitated diffusion involves transport proteins, it is still passive transport because the solute is moving down the concentration gradient.
As was mentioned earlier, small nonpolar molecules can easily diffuse across the cell membrane. However, due to the hydrophobic nature of the lipids that make up cell membranes, polar molecules (such as water) and ions cannot do so. Instead, they diffuse across the membrane through transport proteins. A transport protein completely spans the membrane, and allows certain molecules or ions to diffuse across the membrane. Channel proteins, gated channel proteins, and carrier proteins are three types of transport proteins that are involved in facilitated diffusion.
A channel protein, a type of transport protein, acts like a pore in the membrane that lets water molecules or small ions through quickly. Water channel proteins allow water to diffuse across the membrane at a very fast rate. Ion channel proteins allow ions to diffuse across the membrane.
A gated channel protein is a transport protein that opens a ”gate,” allowing a molecule to pass through the membrane. Gated channels have a binding site that is specific for a given molecule or ion. A stimulus causes the ”gate” to open or shut. The stimulus may be chemical or electrical signals, temperature, or mechanical force, depending on the type of gated channel. For example, the sodium gated channels of a nerve cell are stimulated by a chemical signal which causes them to open and allow sodium ions into the cell. Glucose molecules are too big to diffuse through the plasma membrane easily, so they are moved across the membrane through gated channels. In this way glucose diffuses very quickly across a cell membrane, which is important because many cells depend on glucose for energy.
A carrier protein is a transport protein that is specific for an ion, molecule, or group of substances. Carrier proteins ”carry” the ion or molecule across the membrane by changing shape after the binding of the ion or molecule. Carrier proteins are involved in passive and active transport. A model of a channel protein and carrier proteins is shown in Figure 6.

Figure 6: Facilitated diffusion in cell membrane. Channel proteins and carrier proteins are shown (but not a gated-channel protein). Water molecules and ions move through channel proteins. Other ions or molecules are also carried across the cell membrane by carrier proteins. The ion or molecule binds to the active site of a carrier protein. The carrier protein changes shape, and releases the ion or molecule on the other side of the membrane. The carrier protein then returns to its original shape.
Ion Channels
Ions such as sodium (\(Na^+\)), potassium (\(K^-\)), calcium (\(Ca^{2+}\)), and chloride (\(Cl^-\)), are important for many cell functions. Because they are polar, these ions do not diffuse through the membrane. Instead they move through ion channel proteins where they are protected from the hydrophobic interior of the membrane. Ion channels allow the formation of a concentration gradient between the extracellular fluid and the cytosol. Ion channels are very specific as they allow only certain ions through the cell membrane. Some ion channels are always open, others are ”gated” and can be opened or closed. Gated ion channels can open or close in response to different types of stimuli such as electrical or chemical signals.
Active Transport
In contrast to facilitated diffusion which does not require energy and carries molecules or ions down a concentration gradient, active transport pumps molecules and ions against a concentration gradient. Sometimes an organism needs to transport something against a concentration gradient. The only way this can be done is through active transport which uses energy that is produced by respiration (ATP). In active transport, the particles move across a cell membrane from a lower concentration to a higher concentration. Active transport is the energy-requiring process of pumping molecules and ions across membranes ”uphill” against a gradient.
• The active transport of small molecules or ions across a cell membrane is generally carried out by transport proteins that are found in the membrane.
• Larger molecules such as starch can also be actively transported across the cell membrane by processes called endocytosis and exocytosis.
Sodium-Potassium Pump
Carrier proteins can work with a concentration gradient (passive transport), but some carrier proteins can move solutes against the concentration gradient (from high concentration to low), with energy input from ATP. As in other types of cellular activities, ATP supplies the energy for most active transport. One way ATP powers active transport is by transferring a phosphate group directly to a carrier protein. This may cause the carrier protein to change its shape, which moves the molecule or ion to the other side of the membrane. An example of this type of active transport system, as shown in Figure 7, is the sodium-potassium pump, which exchanges sodium ions for potassium ions across the plasma membrane of animal cells.

Figure 7: The sodium-potassium pump system moves sodium and potassium ions against large concentration gradients. It moves two potassium ions into the cell where potassium levels are high, and pumps three sodium ions out of the cell and into the extracellular fluid.
As is shown in Figure 7, three sodium ions bind with the protein pump inside the cell. The carrier protein then gets energy from ATP and changes shape. In doing so, it pumps the three sodium ions out of the cell. At that point, two potassium ions move in from outside the cell and bind to the protein pump. The sodium-potassium pump is found in the plasma membrane of almost every human cell and is common to all cellular life. It helps maintain cell potential and regulates cellular volume. Cystic fibrosis is a genetic disorder that results in a misshapen chloride ion pump. Chloride levels within the cells are not controlled properly, and the cells produce thick mucus. The chloride ion pump is important for creating sweat, digestive juices, and mucus.
The Electrochemical Gradient
The active transport of ions across the membrane causes an electrical gradient to build up across the plasma membrane. The number of positively charged ions outside the cell is greater than the number of positively charged ions in the cytosol. This results in a relatively negative charge on the inside of the membrane, and a positive charge on the outside. This difference in charges causes a voltage across the membrane. Voltage is electrical potential energy that is caused by a separation of opposite charges, in this case across the membrane. The voltage across a membrane is called membrane potential. Membrane potential is very important for the conduction of electrical impulses along nerve cells.
Because the inside of the cell is negative compared to outside the cell, the membrane potential favors the movement of positively charged ions (cations) into the cell, and the movement of negative ions (anions) out of the cell. So, there are two forces that drive the diffusion of ions across the plasma membrane—a chemical force (the ions’ concentration gradient), and an electrical force (the effect of the membrane potential on the ions’ movement). These two forces working together are called an electrochemical gradient.
Vesicles and Active Transport
Some molecules or particles are just too large to pass through the plasma membrane or to move through a transport protein. So cells use two other methods to move these macromolecules (large molecules) into or out of the cell. Vesicles or other bodies in the cytoplasm move macromolecules or large particles across the plasma membrane. There are two types of vesicle transport, endocytosis and exocytosis.
Endocytosis and Exocytosis
Endocytosis is the process of capturing a substance or particle from outside the cell by engulfing it with the cell membrane. The membrane folds over the substance and it becomes completely enclosed by the membrane. At this point a membrane-bound sac, or vesicle pinches off and moves the substance into the cytosol. There are two main kinds of endocytosis:
• Phagocytosis or ”cellular eating,” occurs when the dissolved materials enter the cell. The plasma membrane engulfs the solid material, forming a phagocytic vesicle.
• Pinocytosis or ”cellular drinking,” occurs when the plasma membrane folds inward to form a channel allowing dissolved substances to enter the cell, as shown in Figure 8. When the channel is closed, the liquid is encircled within a pinocytic vesicle.

Figure 8: Transmission electron microscope image of brain tissue that shows pinocytotic vesicles. Pinocytosis is a type of endocytosis.
Exocytosis describes the process of vesicles fusing with the plasma membrane and releasing their contents to the outside of the cell, as shown in Figure 9. Exocytosis occurs when a cell produces substances for export, such as a protein, or when the cell is getting rid of a waste product or a toxin. Newly made membrane proteins and membrane lipids are moved on top the plasma membrane by exocytosis.

Figure 9: Mode of exocytosis at a synaptic junction, where two nerve cells meet. Chemical signal molecules are released from nerve cell A by exocytosis, and move toward receptors in nerve cell B. Exocytosis is an important part in cell signaling.
Homeostasis and Cell Function
Homeostasis refers to the balance, or equilibrium within the cell or a body. It is an organism’s ability to keep a constant internal environment. Keeping a stable internal environment requires constant adjustments as conditions change inside and outside the cell. The adjusting of systems within a cell is called homeostatic regulation. Because the internal and external environments of a cell are constantly changing, adjustments must be made continuously to stay at or near the set point (the normal level or range). Homeostasis is a dynamic equilibrium rather than an unchanging state. The cellular processes discussed all play an important role in homeostatic regulation.
Cell Communication
To survive and grow, cells need to be able to ”talk” with their cell neighbors and be able to detect change in their environment. Talking with neighbors is even more important to a cell if it is part of a multicellular organism. The billions of cells that make up your body need to be able to communicate with each other to allow your body to grow, and to keep you alive and healthy. The same is true for any organism. Cell signaling is a major area of research in biology today. Recently scientists have discovered that many different cell types, from bacteria to plants, use similar types of communication pathways, or cellsignaling mechanisms. This suggests that cell-signaling mechanisms evolved long before the first multicellular organism did.
The Language of Cells
For cells to be able to signal to each other, a few things are needed:
• a signal
• a cell receptor, which is usually on the plasma membrane, but can be found inside the cell
• a response to the signal
Cells that are communicating may be right next to each other or far apart. The type of chemical signal a cell will send differs depending on the distance the message needs to go. For example, hormones, ions, and neurotransmitters are all types of signals that are sent depending on the distance the message needs to go.
The target cell then needs to be able to recognize the signal. Chemical signals are received by the target cell on receptor proteins. As discussed earlier, most receptor proteins are found in the plasma membrane. Most receptors proteins are found on the plasma membrane, but some are also found inside the cell. These receptor proteins are very specific for only one particular signal molecule, much like a lock that recognizes only one key. Therefore, a cell has lots of receptor proteins to recognize the large number of cell signal molecules. There are three stages to sending and receiving a cell ”message:” reception, transduction, and response.
Signal Receptors
Cell-surface receptors are integral proteins—they reach right through the lipid bilayer, spanning from the outside to the inside of the cell. These receptor proteins are specific for just one kind of signal molecule. The signaling molecule acts as a ligand when it binds to a receptor protein. A ligand is a small molecule that binds to a larger molecule. Signal molecule binding causes the receptor protein to change its shape. At this point the receptor protein can interact with another molecule. The ligand (signal molecule) itself does not pass through the plasma membrane.
In eukaryotic cells, most of the intracellular proteins that are activated by a ligand binding to a receptor protein are enzymes. Receptor proteins are named after the type of enzyme that they interact with inside the cell. These enzymes include G proteins and protein kinases, likewise there are G-protein-linked receptors and tyrosine kinase receptors. A kinase is a protein involved in phosphorylation. A G-protein linked receptor is a receptor that works with the help of a protein called a G-protein. A G-protein gets its name from the molecule to which it is attached, guanosine triphosphate (GTP), or guanosine diphosphate (GDP). The GTP molecule is similar to ATP.
Once G proteins or protein kinase enzymes are activated by a receptor protein, they create molecules called second messengers. A second messenger is a small molecule that starts a change inside a cell in response to the binding of a specific signal to a receptor protein. Some second messenger molecules include small molecules called cyclic nucleotides, such as cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP). Calcium ions (\(Ca^{2+}\)) also act as secondary messengers. Secondary messengers are a part of signal transduction pathways.
Signal Transduction
A signal-transduction pathway is the signaling mechanism by which a cell changes a signal on it surface into a specific response inside the cell. It most often involves an ordered sequence of chemical reactions inside the cell which is carried out by enzymes and other molecules. In many signal transduction processes, the number of proteins and other molecules participating in these events increases as the process progresses from the binding of the signal. A ”signal cascade” begins. Think of a signal cascade as a chemical domino-effect inside the cell, in which one domino knocks over two dominos, which in turn knock over four dominos, and so on. The advantage of this type of signaling to the cell is that the message from one little signal molecule can be greatly amplified and have a dramatic effect.
G protein-linked receptors are only found in higher eukaryotes, including yeast, plants, and animals. Your senses of sight and smell are dependent on G-protein linked receptors. The ligands that bind to these receptors include light-sensitive compounds, odors, hormones, and neurotransmitters. The ligands for G-protein linked receptors come in different sizes, from small molecules to large proteins. G protein-coupled receptors are involved in many diseases, but are also the target of around half of all modern medicinal drugs.
The process of how a G-protein linked receptor works is outlined in Figure 10.

Figure 10: How a G-protein linked receptor works with the help of a G-protein. In panel C, the second messenger cAMP can be seen moving away from the enzyme.
| A. | A ligand such as a hormone (small, purple molecule) binds to the G-linked receptor (red molecule). Before ligand binding, the inactive G-protein (yellow molecule) has GDP bound to it. |
| B. | The receptor changes shape and activates the G-protein and a molecule of GTP replaces the GDP. |
| C. | The G-protein moves across the membrane then binds to and activates the enzyme (green molecule). This then triggers the next step in the pathway to the cell’s response. After activating the enzyme, the Gprotein returns to its original position. The second messenger of this signal transduction is cAMP, as shown in C. |
The sensing of the external and internal environments at the cellular level relies on signal transduction. Defects in signal transduction pathways can contribute or lead to many diseases, including cancer and heart disease. This highlights the importance of signal transductions to biology and medicine.
Signal Responses
In response to a signal, a cell may change activities in the cytoplasm or in the nucleus that include the switching on or off of genes. Changes in metabolism, continued growth, movement, or death are some of the cellular responses to signals that require signal transduction.
Gene activation leads to other effects, since the protein products of many of the responding genes include enzymes and factors that increase gene expression. Gene expression factors produced as a result of a cascade can turn on even more genes. Therefore one stimulus can trigger the expression of many genes, and this in turn can lead to the activation of many complex events. In a multicellular organism these events include the increased uptake of glucose from the blood stream (stimulated by insulin), and the movement of neutrophils to sites of infection (stimulated by bacterial products). The set of genes and the order in which they are activated in response to stimuli are often called a genetic program.
Images courtesy of:
http://en.wikipedia.org/wiki/Image:Semipermeable_membrane.png. Public Domain.
Mariana Ruiz. http://commons.wikimedia.org/wiki/File: Scheme_simple_diffusion_in_cell_membrane-en.svg. Public Domain.
http://en.wikipedia.org/wiki/Image: Turgor_pressure_on_plant_cells_diagram.svg. Public Domain.
Mnolf. http://en.wikipedia.org/wiki/Image:Rhoeo_Discolor_-_Plasmolysis.jpg. GNU-FDL & CC-BY-SA.
Jasper Nance. CC-BY-SA.
Mariana Ruiz. http://commons.wikimedia.org/wiki/File: Scheme_facilitated_diffusion_in_cell_membrane-en.svg. Public Domain.
Mariana Ruiz. http://commons.wikimedia.org/wiki/File: Scheme_sodium-potassium_pump-en.svg. Public Domain.
Louisa Howard, Miguel Marin-Padilla. Public Domain.
Dake. http://en.wikipedia.org/wiki/Image:Synapse_diag1.png. GNU-FDL.
Bensaccount. [Retrieved and modified from http://en.wikipedia.org/wiki/Image:GPCR_mechanism.png ]. Public Domain.